色素体/葉緑体の成立と多様性 |
||||
執筆:鈴木雅大・大田修平 作成日:2010年10月5日(2022年9月2日更新) |
||||
 |
||||
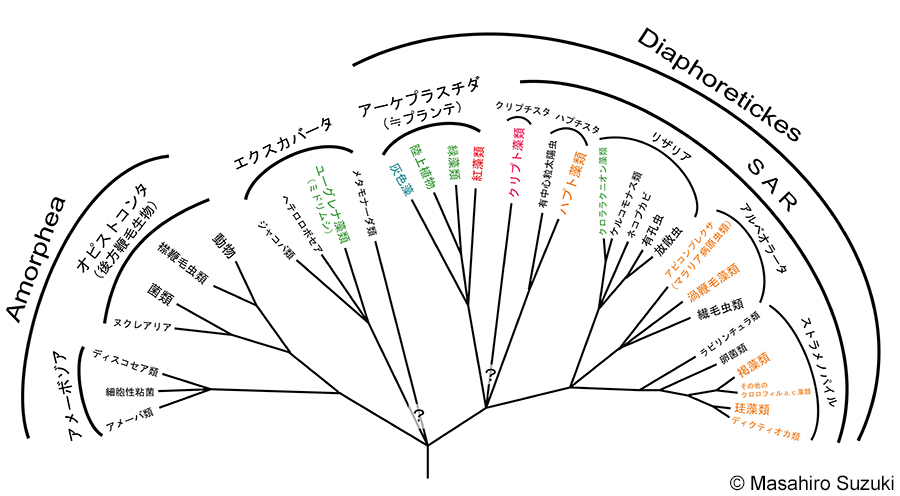
| 酸素発生型光合成を行う生物を「植物」と呼ぶとするならば,生物の世界の中で「植物」はなんと多様な事でしょう。上の図は,Adl et al. (2019)を参考に作図したもので,真核生物の主なスーパーグループを示したものです(前回の更新(Adl et al. 2012)で作成した図はこちら)。各スーパーグループの系統的位置やグループ名は近年の仮説とは若干異なりますが,真核生物の世界にどのような生き物がいるかを示すのに分かりやすいことからこの図を用いました。色素体/葉緑体を持ち,光合成を行う真核生物をカラーで示しました。緑,赤,青,褐色など様々な色があるのは後述する色素体/葉緑体の形態の多様性に関係しています。この図を見ると,光合成を行う真核生物は,オピストコンタ(動物,菌類),アメーボゾアを除く全ての系統群に存在していることが分かると思います(注1)。光合成を行うオルガネラ(細胞小器官)は色素体です。現在,色素体がただ一度の細胞内共生に起源を持つという考えは,広く受け入れられています(注2)。その色素体が真核生物の様々な系統群で見られるのはどういう事なのでしょうか。光合成真核生物の多様性を学ぶには,色素体がどのようにして獲得されてきたのかを理解する必要があります。皆さんの中には,高等学校の教科書で,色素体(葉緑体)やミトコンドリアは,真核生物が原核生物を細胞内共生(endosymbiosis)によってオルガネラとしたという「細胞内共生説」を習ったことがある方もおられるでしょう。細胞内共生とは,ある真核生物が他の生物を食作用によって細胞内に取り込み,これを共生体として維持,最終的にオルガネラとする現象です。ミトコンドリアと葉緑体はその最たる例です。ミトコンドリアは真核生物がα-プロテオバクテリア(現生のリケッチアに近縁と考えられている)を細胞内共生したものですし,緑色植物の葉緑体はシアノバクテリアの仲間を細胞内共生したものです。このコーナーでは細胞内共生による色素体の獲得と色素体の多様性について解説します。
注1: ミドリガイの仲間(軟体動物,ウミウシの仲間)には,クレプトクロロプラスト(盗葉緑体)によって光合成を行うものが知られていますが,色素体として成立したものではありません。 |
||||
| 葉緑体か色素体か | ||||
| 光合成を行うオルガネラ(細胞小器官)のことを「葉緑体 chloroplast」または「色素体 plastid」といいます。私達が生物教育で最初に習う,教えるテーマは「細胞」で,「葉緑体」という名称及び機能は多くの方が知っていると思います。しかし,厳密に言うと「葉緑体」という名称は,緑色をした色素体を指し,分類群では緑色植物のみに用いられます。後述する通り,光合成生物は多様で,"葉緑体"の種類(色,構造,含有色素など)は様々です。緑色植物が持つ「葉緑体」以外は「色素体」と呼んでいます。緑色植物でも,貯蔵デンプンの前駆体のような光合成色素を持たない"葉緑体"を白色体(leucoplast)と呼びます。日本人にとって「葉緑体」という言葉は広く受け入れられているので,一般的に「色素体」も含めて「葉緑体」と呼ぶことが多いですが,本稿では光合成生物の多様性を理解するため,「葉緑体」と「色素体」を厳密に区別して用いています。 | ||||
| 一次細胞内共生(一次共生)と一次植物 | ||||
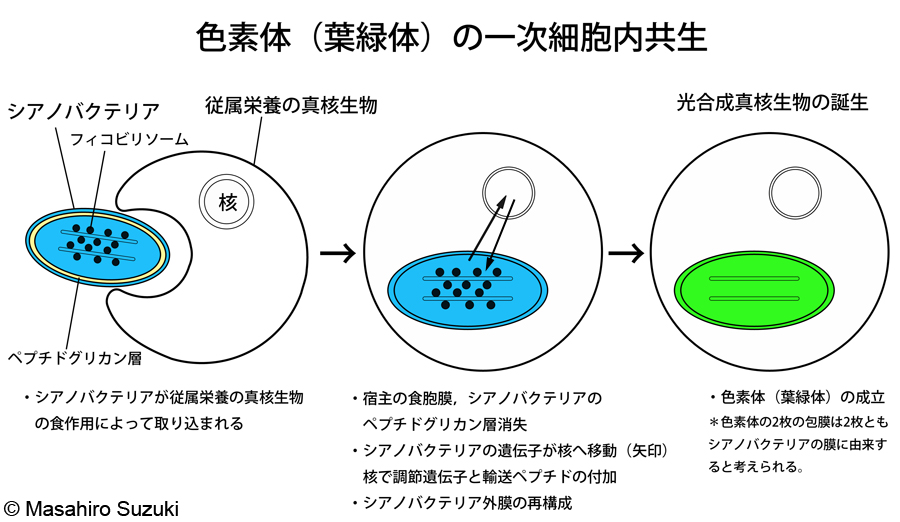
| 従属栄養の真核生物がシアノバクテリア(正しくはシアノバクテリア様の祖先生物)を細胞内共生によって取り込み,色素体とする事を一次細胞内共生(一次共生)といい,一次共生によって誕生した光合成真核生物を一次植物といいます。現在の真核生物の分類体系では,一次植物をスーパーグループ アーケプラスチダ(=「最初の植物」,「真の植物」)として定義しています。 | ||||
 |
||||
| 一次共生の模式図:従属栄養の真核生物がシアノバクテリアを食作用によって取り込みます。次に取り込んだ宿主の食胞膜と取り込まれたシアノバクテリアのペプチドグリカン層が消失します。シアノバクテリアの遺伝子が宿主の核に移動します。調節遺伝子と輸送ペプチドを付加され,核コードの色素体遺伝子となります。こうして核に移動した色素体関連遺伝子は核でmRNAに転写された後,細胞質で翻訳され,色素体タンパク質となります。このタンパク質は輸送ペプチドによって色素体内へ輸送されます。シアノバクテリアの外膜は,核から運ばれてきた色素体タンパク質を色素体内に輸送するために再構成されます。こうして現生の色素体が誕生したと考えられています。 | ||||
| 余談:色素体の膜の由来について | ||||
| 著者は高校時代,色素体の外側の膜は宿主の食胞膜であると教わりました。Margulisの細胞内共生説の肝であり,食べた宿主の膜が残っているということで,説得力のある仮説だったと思います。けれども,上述した色素体へのタンパク質輸送機構の研究の進展により,近年この説はほぼ否定され,色素体の2枚の包膜は2枚ともシアノバクテリアの膜に由来していると考えられています。色素体と核,細胞質との物質の移動を考えれば,元々ある膜の輸送チャネルなどを再利用した方が最節約的であり,むしろ2枚の包膜が共にシアノバクテリアに由来していなければ不自然です。ただし,外膜と内膜のタンパク質構成が完全に解明された訳ではありません。色素体の分裂に関わるタンパク質の中には,共生後に宿主側から付加されたと考えられるものもあり,新たに獲得された膜タンパク質も少なくないように思われます(宮城島 2014)。細胞内共生説を巡っては,これまでも仮説が二転三転しており,宿主の食胞膜説が復権する可能性も無くはありません。このような興味の尽きない難解さと不確定さが生物学の特殊な点と面白い点ではないかと思っています。 | ||||
| 一次植物の仲間 | ||||
| 緑色植物,紅藻植物,灰色藻植物の3つの分類・系統群が知られています。いずれも色素体(葉緑体)包膜が2重です。 | ||||
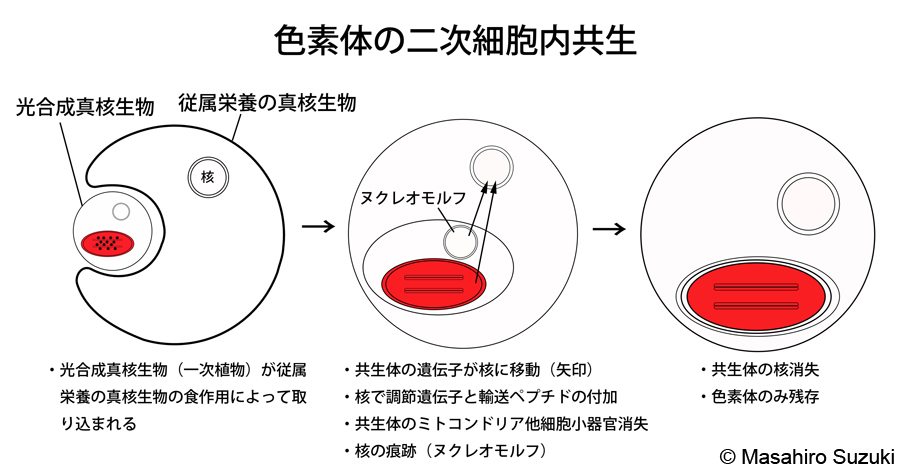
| 二次細胞内共生(二次共生)と二次光合成生物(二次植物) | ||||
| 従属栄養の真核生物が光合成真核生物,すなわち一次植物を細胞内共生によって取り込み,色素体とする事を二次細胞内共生(二次共生)といいます。「二次植物」と呼ばれることもありますが,二次共生によって色素体を獲得した光合成生物は「真の植物」として定義されている一次植物とは系統的に異なることから,「植物」と呼ばず,「二次光合成生物」と呼んでいます。 | ||||
 |
||||
| 二次光合成生物(二次植物)の仲間 | ||||
| 二次光合成生物(二次植物)は,大きく二つに分けられます。緑色植物を取り込み,主要な光合成色素にクロロフィルa, bを持つ真核藻類と,紅藻植物を取り込み,主要な光合成色素にクロロフィルaと新たに獲得したクロロフィルcを持つ真核藻類です。注目されるのは,中央の図に示したヌクレオモルフです。これは真核生物が真核生物を細胞内共生した大きな証拠で,共生体の痕跡的な核と考えられています。さらに共生が進むとヌクレオモルフは消失し,色素体のみが残されると推定されます。ヌクレオモルフを持っている二次光合成生物がクロララクニオン藻類とクリプト藻類で,色素体の成立の途中段階にあると考えられる生物が現生にも生息していた事が,我々が真核生物の進化・系統を読み解く上での重要なヒントとなっています。 | ||||
| 緑色植物を細胞内共生した二次光合成生物…クロララクニオン藻類,ミドリムシ(ユーグレナ)藻類 | ||||
| 紅藻植物を細胞内共生した二次光合成生物…クリプト藻類
注.従来,紅藻植物を細胞内共生した二次光合成生物は,クリプト藻類,ハプト藻類,黄色藻類(不等毛藻類),渦鞭毛類と考えられてきましたが,近年,紅藻植物を細胞内共生したのは,クリプト藻類のみで,他のグループは,クリプト藻類,ハプト藻類,黄色藻類などを細胞内共生したとする仮説が提唱されています(Bodył et al. 2009, Strassert et al. 2021)。 |
||||
| 三次細胞内共生,四次細胞内共生 | ||||
| 緑色植物や紅藻植物ではなく,二次光合成生物であるクリプト藻,ハプト藻,黄色藻類を細胞内共生して色素体とする現象です。一部の渦鞭毛藻類で確認されていたものですが,近年,クリプト藻類以外のクロロフィルcを持つ真核藻類は,クリプト藻類,ハプト藻類,黄色藻類などを細胞内共生したとする仮説が提唱されています(Bodył et al. 2009, Strassert et al. 2021)。回数でいえば「三次」,「四次」となりますが,ここまで複雑になると「三次」,「四次」と区別するのは,かえって分かりにくくなるように思えます。本サイトでは,一次細胞内共生と二次細胞内共生までは,区別しますが,「三次」,「四次」については省略しました。 | ||||
| 色素体(葉緑体)の形態の多様性 | ||||
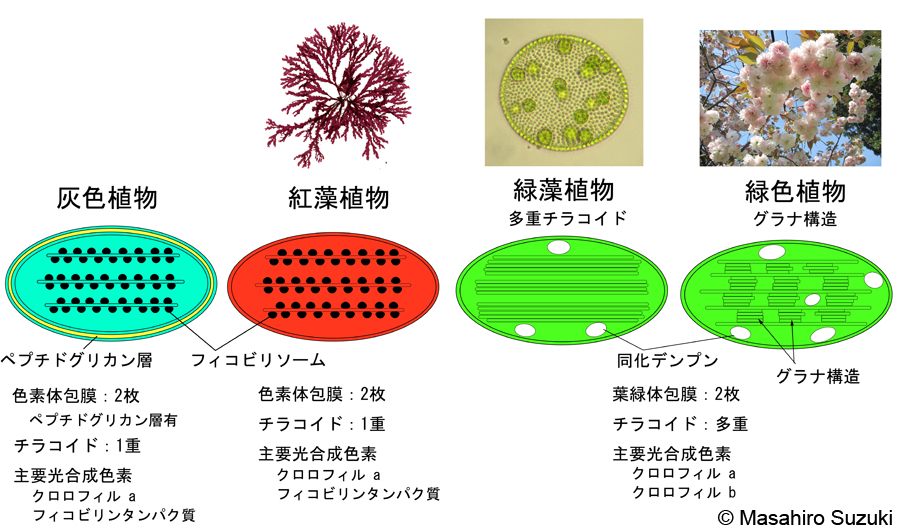
| 色素体(葉緑体)の形態は,色素体包膜の枚数,チラコイド,光合成色素の違いなどによっていくつかのタイプに分けられます。一次植物の色素体/葉緑体包膜は例外なく2枚ですが,二次光合成生物の色素体包膜は3枚以上です。これは上述の一次共生と二次共生と関係があると考えられています。ここでは,光合成真核生物に見られる主な色素体/葉緑体の形態を図と共に紹介します。 | ||||
| 一次植物の色素体/葉緑体 | ||||
 |
||||
| 灰色植物の色素体:灰色植物の色素体には,シアノバクテリアと同じく2枚の膜の間にペプチドグリカン層が見られます。また,図には示していませんが,色素体の中央部にシアノバクテリアに見られるカルボキシソームを持っています。このため,以前は色素体ではなく細胞内に共生したシアノバクテリアだと考えられ,「シアネル」と呼ばれていました。現在では「シアネル」の全ゲノムが解読され,細胞内共生シアノバクテリアではなく,色素体として成立した段階にあると考えられています。 | ||||
| 紅藻植物の色素体:2枚の膜に包まれ,チラコイドは1枚ずつ独立しています。チラコイドの表面にフィコビリソームという構造が付着しています。フィコビリソームには主要なアンテナ色素系を担うフィコビリンタンパク質が組み込まれています。 | ||||
| 緑色植物の葉緑体:高等学校の生物の教科書に載っている色素体で,厳密に言えばこのグループの色素体のみを「葉緑体」と呼びます。多重チラコイドを持ちます。一部の緑藻植物,コケ植物と維管束植物(シダ・裸子・被子植物を含む)の葉緑体はグラナ構造を作ります。光合成産物の代表的な貯蔵物質であるデンプンが葉緑体の内部に蓄積されるのは緑色植物のみで,緑色植物の大きな特徴の一つです。 | ||||
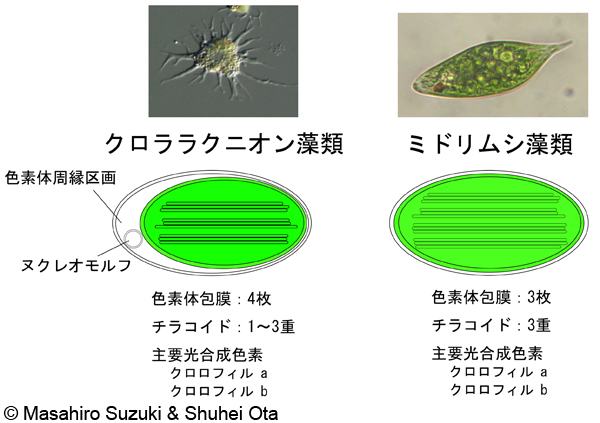
| 光合成生物の色素体 1. クロララクニオン藻類とミドリムシ藻類 | ||||
 |
||||
| 共に一次植物の緑色植物(緑藻類)を取り込んで成立した色素体(*)で,主要光合成色素としてクロロフィルaとbを持っています。*:緑色植物に由来するので「葉緑体」と呼んでも差し支えはないと思いますが,一次植物と明確に区別したいことから,本稿では「色素体」としました。 | ||||
| クロララクニオン藻類の色素体:4枚の膜に包まれています。外側の2枚と内側の2枚の色素体包膜との間に色素体周縁区画と呼ばれる空間があります。この空間にヌクレオモルフという共生体の核の痕跡と考えられる構造が見られます。チラコイドは普通3重ですが,1重,2重のものも時々見られます。クロララクニオン藻類の形態・分類・進化で更に詳しい解説を読むことが出来ます。 | ||||
ミドリムシ(ユーグレナ)藻類の色素体:3枚の膜に包まれています(注)。 注:2009年の日本植物学会(山形大会)で,奈良女子大のグループがミドリムシの色素体を加圧凍結固定法(できるだけ生きたままの状態を反映する固定法)により電子顕微鏡観察したところ,色素体包膜が4枚観察されたことが報告されました。この研究のさらなる展開に期待したいところです。 |
||||
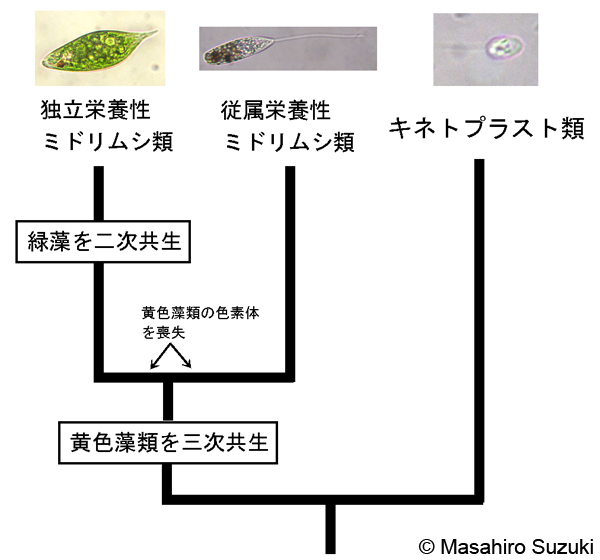
| ミドリムシ藻類で明らかになった色素体の入れ替え(連続的二次細胞内共生) | ||||
| ミドリムシ藻類の色素体は,上述の通り緑色でクロロフィルaとbを持ち,緑藻を二次共生したものと考えられてきましたが,Maruyama et al. (2011)とYang et al. (2011) により,ミドリムシの祖先は,珪藻に近い黄色藻類を細胞内共生していたことが示唆されました(むかしむかし、ミドリムシは紅かった?)。珪藻は,スーパーグループ SARに所属する黄色藻類で,紅藻を二次共生したグループです。ややこしいことですが,ミドリムシの祖先は,珪藻のような黄色藻類を三次共生した後,一度獲得した色素体を捨て,新たに緑藻を二次共生したということになります。すなわち「色素体の入れ替え」を行ったと考えられます。同様の現象は,渦鞭毛藻類(SAR,アルベオラータ)でも知られており,二次共生あるいは三次共生はこれまで考えられてきた以上に複雑かつ高頻度で起こった現象のようです。なお,著者の素朴な疑問ですが,黄色藻類を細胞内共生したのならば,紅ではなく黄色だったのではないでしょうか? | ||||
 |
||||
| ミドリムシ(ユーグレナ)門の系統と色素体獲得の模式図 | ||||
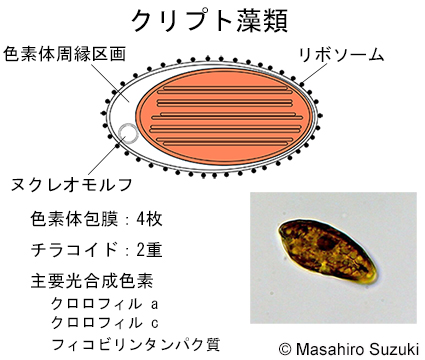
| 光合成生物の色素体 2. クリプト藻類 | ||||
 |
||||
| クリプト藻類の色素体は,紅藻植物を細胞内共生して成立したと考えられ,4枚の膜で包まれています。外側の2膜の内,最外膜の表面にはリボソームが付着し,この膜は核の外膜と連結していて粗面小胞体の形態を取ります。このため,4枚の色素体包膜の外側の2枚を色素体ER(小胞体)と呼んでいます(注)。クロララクニオン藻類と同様に色素体周縁区画にヌクレモルフが見られます。光合成真核生物の中で唯一2重チラコイドが見られます。アンテナ色素として新たにクロロフィル cを持っています。紅藻植物のアンテナ色素であるフィコビリンタンパク質を残していますが,紅藻植物とは違い,フィコビリンタンパク質はチラコイド内腔に組み込まれているためフィコビリソームは作りません。
注:色素体ERにリボソームが付着している生物学的理由は不明です。 |
||||
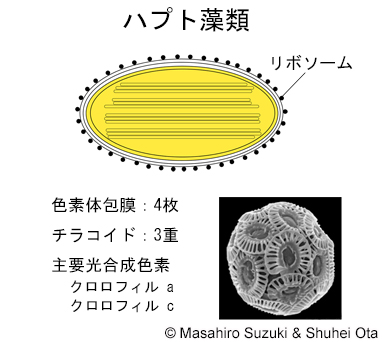
| 光合成生物の色素体 3. ハプト藻類 | ||||
 |
||||
| ハプト藻類の色素体は,紅藻植物を細胞内共生して成立したと考えられてきましたが,近年は,クリプト藻類または黄色藻類(不等毛藻類)を細胞内共生したとする仮説が提唱されています(Bodył et al. 2009, Strassert et al. 2021)。色素体は4枚の膜に包まれ,外側の2膜の内,最外膜の表面にはリボソームが付着し,色素体ER(小胞体)となっています。チラコイドは3重です。 | ||||
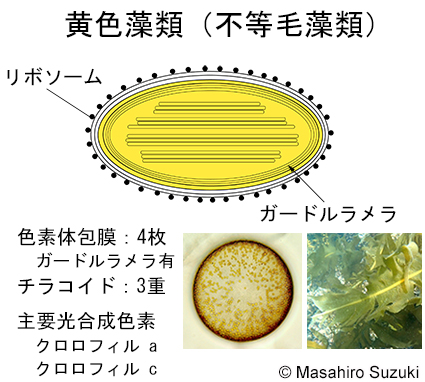
| 光合成生物の色素体 4. 黄色藻類(不等毛藻類) | ||||
 |
||||
| 黄色藻類(不等毛藻類)の色素体は,紅藻植物を細胞内共生して成立したと考えられてきましたが,近年は,クリプト藻類を細胞内共生したとする仮説が提唱されています(Bodył et al. 2009, Strassert et al. 2021)。色素体は4枚の膜に包まれ,外側の2膜の内,最外膜の表面にはリボソームが付着し,色素体ER(小胞体)となっています。色素体包膜の直下にガードルラメラという袋状のラメラがあり,その中に三重のチラコイドが入っています。一部の黄色藻類には,ガードルラメラを持たないものがあります。 | ||||
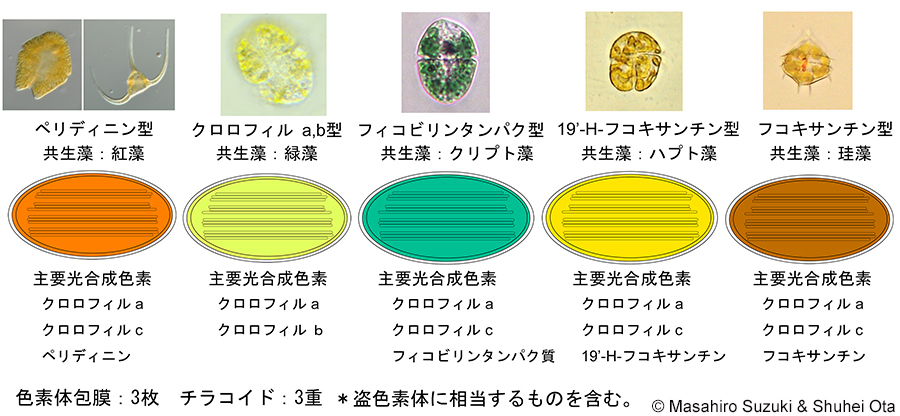
| 光合成生物の色素体 5. 渦鞭毛類 | ||||
 |
||||
| 渦鞭毛類は藻類といいながら全体の半分位は色素体を持たない従属栄養性の単細胞生物で,渦鞭毛虫類とも呼ばれています。この仲間は,他の生物と共生関係を持ちやすいようで,他の二次光合成生物が紅藻植物あるいは緑色植物(緑藻)のいずれかを共生体としているのに対し,この仲間は分かっているだけで紅藻植物,緑色植物,クリプト藻類,ハプト藻類,黄色藻類(珪藻)の5つのグループを細胞内共生しています。
従来,渦鞭毛類の色素体は紅藻植物を細胞内共生して成立したと考えられてきましたが,近年は,ハプト藻類あるいは黄色藻類を細胞内共生したという仮説が提唱されています(Bodył et al. 2009, Strassert et al. 2021)。ハプト藻類の色素体は,クリプト藻類または黄色藻類を細胞内共生したものと考えられ,黄色藻類の色素体はクリプト藻類を細胞内共生たと考えられることから,渦鞭毛藻類の色素体は,回数で言えば「四次」細胞内共生ということになります。 渦鞭毛藻類の色素体は3枚の膜で包まれていますが,クロメラとアピコンプレクサの色素体は4枚の膜で包まれていることから,渦鞭毛類の祖先生物の色素体はかつて4枚の膜に包まれていて,それが進化の過程で1枚失われたと考えられます。4枚の膜のうちどれが消失し,また,なぜ消失したのでしょうか?現在のところ,これに関する明確な答えは出ていません。今後の細胞生物学的研究,特に色素体タンパク質の輸送の研究アプローチにより,解決できるかもしれません。 渦鞭毛類の中では少数派ですが,ペリディニン型以外の仲間の色素体はどのようにして獲得されたのでしょうか。渦鞭毛類の祖先生物は紅藻植物由来の色素体を持っていたと考えられることから,緑色植物を細胞内共生したもの(クロロフィルa, b型)は,紅藻物由来の色素体を一度失い,緑色植物由来の色素体を置き換えた事になります。これを連続的二次細胞内共生と呼んでいます。残りの3型(フィコビリンタンパク型,19'-H-フコキサンチン型,フコキサンチン型)は,それぞれ紅藻植物由来の色素体を失った後で,二次光合成生物であるクリプト藻類,ハプト藻類,珪藻を細胞内共生したものなので三次細胞内共生(注2)と呼んでいます。渦鞭毛藻類にはここに挙げた以外に,細胞内共生して色素体が成立する途上の段階にあると考えられる種類が知られています(注2)。このように渦鞭毛類は共生について言えば極めて多様なグループです。なぜこのグループだけがここまで多様化しているのか明確な答えは出ていません。 注1: ある従属栄養性真核生物が二次光合成生物を共生体としたのならば純粋に三次細胞内共生と呼んで問題ないと思いますが,一度色素体を失った後に色素体を置き換えた場合,連続的二次共生のような特別な言い方をした方が良いかもしれません。 注2: 緑色植物を連続的二次細胞内共生したクロロフィルa, b型の色素体,珪藻を三次細胞内共生したフコキサンチン型の色素体は,実際には色素体を成立する前の段階(段階2または3に相当)と考えられます。クリプト藻類を三次共生したフィコビリンタンパク型の色素体を持つものには,色素体を成立する前の段階(段階2)に相当するものとクレプトプラスチド(盗色素体)段階に相当するものがあり,後者については通常色素体とは言いません。すなわち,渦鞭毛類の色素体として挙げた5型の内,色素体と成立した段階とはっきりと言えるのはペリディニン型(共生体は紅藻植物)と19'-H-フコキサンチン型(共生体はハプト藻類)のみです。クロロフィルa, b型(共生体は緑色植物)は共生体の核が残存していますが,色素体として機能していると考えられます。フコキサンチン型(共生体は珪藻)とフィコビリンタンパク型(共生体はクリプト藻類)の2型については色素体と呼んで良いかどうか検討中です。 余談になりますが,渦鞭毛類の中には,横溝の中にシアノバクテリアを細胞外共生させているものや,Symbiodiniumのようにズーザンテラとして渦鞭毛類自身がサンゴなどの動物の共生藻となっているものもあります。これらは共生ではありますが,細胞内共生として共生体を色素体化するものとは便宜上分けて考えています。前者については細胞外共生とはいえ,渦鞭毛類にとっては細胞内共生と変わらないのかもしれません。渦鞭毛類を見ていると生物にとっての「共生」とは何なのか考えさせられます。 |
||||
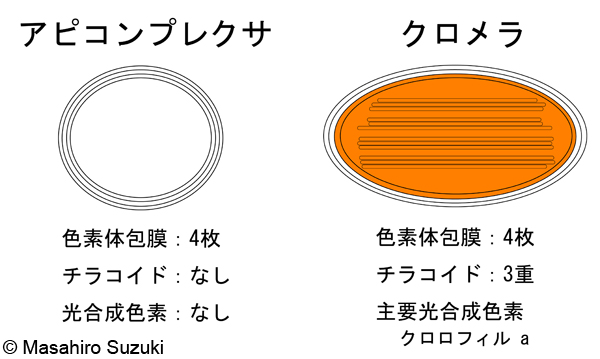
| 光合成生物の色素体 6. アピコンプレクサとクロメラ | ||||
 |
||||
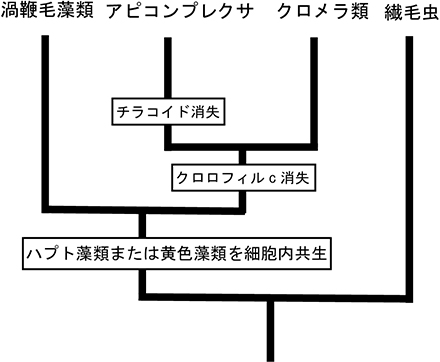
| アピコンプレクサとクロメラは,上述の渦鞭毛類,ゾウリムシやツリガネムシ,ラッパムシを含む繊毛虫類と共にアルベオラータ上門に所属しています。アピコンプレクサというのは聞きなれない名前かもしれませんが,ここに所属しているマラリア原虫(Plasmodium)やトキソプラズマ症原虫(Toxoplasma)は,蚊が媒介して感染し,2005年度の統計では世界で年間3-5億人もの感染者と100-150万人の死亡者を出しているマラリアという病気の病原体です。人類にとって極めて恐ろしい病原生物ですが,細胞内にアピコプラストという退化した色素体を持っており,藻類・プロチストの分類・進化を語るには欠かせないグループです。アピコプラストは4枚の色素体包膜で包まれ,チラコイドや光合成色素は失われています。光合成は行いませんが,脂肪酸の代謝などを担っています。4枚の膜に包まれることから二次共生によって獲得された色素体であること,アピコプラストにコードされた遺伝子解析から共生体は紅藻植物と考えられています(注)。
2008年2月,イギリスの科学雑誌ネイチャー(Nature)にクロメラ(Chromela velia)という「藻類」が記載されました(Moore et al. 2008)。この生物は遺伝子解析や微細構造解析よりアピコンプレクサ類に近縁である(系統樹ではアピコンプレクサと姉妹関係になる)ことが分かり,渦鞭毛藻類とアピコンプレクサ類をつなぐ生物として注目されています。サンゴの一種に共生する藻類で,C. velia一種しか知られていませんが,既知のどの門にも属さない新門,クロメラ門のメンバーとして記載されました。遺伝子解析から渦鞭毛藻類,アピコンプレクサ,クロメラの祖先生物が紅藻植物を二次共生した事が示唆されました(注)。クロメラの色素体は4枚の膜で包まれ,チラコイドは3重です。光合成色素はクロロフィル aのみで,クロロフィル cはクロメラとおそらくアピコンプレクサの共通祖先において失われたと考えられます。 注.近年は,アピコンプレクサとクロメラの色素体も,渦鞭毛藻類と同様にハプト藻類あるいは黄色藻類を細胞内共生したものと考えられています(Strassert et al. 2021)。 |
||||
 |
 |
|||
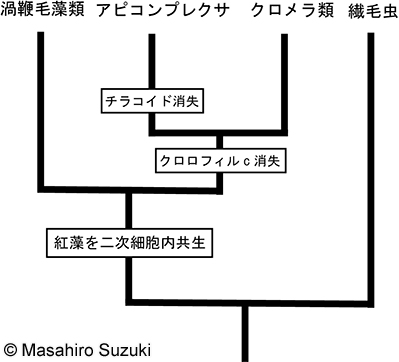
| アルベオラータ上門の系統と色素体の獲得の模式図(左図:紅藻類を二次細胞内共生したとする従来の仮説。右図:ハプト藻類または黄色藻類を細胞内共生したという仮説) | ||||
| 二次細胞内共生による"植物化"の過程 | ||||
| 井上(2007)は,二次光合成生物が色素体を獲得するまでを4つの段階に分けています。共生体が色素体となるまでの過程は,共生体が宿主によって制御されていく過程と言い換える事が出来ます。 | ||||
| 1. クレプトプラスチド(盗色素体)段階 | ||||
| 捕食した共生体(真核光合成生物)をある程度細胞内に保持して光合成させ,何ヶ月も利用し続けます。捕食によって共生体を常に補充する必要があります。渦鞭毛類の一部(Amphidinium latum, Gymnodinium acidotum,Nusuttodinium aeruginosumなど)と繊毛虫の一部(ミドリゾウリムシなど)でこの段階にある生物が見つかっています。 | ||||
| 共生体の状態:一定期間消化されない,オルガネラは大部分残存,宿主と共生体の分裂は同調していません。 | ||||
| 2. 共生体の継続的保持 | ||||
| 共生体が宿主の分裂と共に分裂するため,新しい共生体を補充する必要がありません。この時点で「光合成生物になった」と言う事が出来ます。渦鞭毛類のDurinskia balticaやPeridinium foliaceumなどがこの段階にあると考えられています。 | ||||
| 共生体の状態:核とミトコンドリアを除くオルガネラの消失。分裂が制御され,宿主の細胞周期と同調しています。 | ||||
| 3. 共生体遺伝子の核への移行 | ||||
| クリプト藻類,クロララクニオン藻類,渦鞭毛類の一部(Lepidodinium viride,Gymnodinium chlorophorumなど)がこの段階にあります。二次光合成生物の色素体が成立する直前の段階と考えられています。 | ||||
| 共生体の状態:ミトコンドリア消失,核の痕跡(ヌクレオモルフ)が見られます。 | ||||
| 4. 色素体の成立 | ||||
| 共生体の核が完全に消失し,色素体のみが残された段階です。 | ||||
| クレプトプラスチド(盗色素体)現象 Kleptoplasty | ||||
| 従属栄養性の真核生物が光合成真核生物,すなわち藻類を細胞内に取り込み,色素体/葉緑体として利用する現象です。ミドリガイの仲間のウミウシのように,緑藻を食べてその葉緑体を細胞内に保持する場合は,クレプトクロロプラスト(盗葉緑体,Kleptochloroplast)と呼びます。クレプトプラスチドには幾つかのタイプがあり,上述の色素体成立の第1段階に相当すると考えられるものもあれば,この先第2段階に進むとは考えにくい,色素体の成立とは関係が無いかもしれないものもあります。改めて本HPのコラムとして解説したいと思います。 | ||||
| ハテナの発見 | ||||
| 大系統を勉強するにはや用語解説のコーナーでも紹介していますが,ハテナがこんなにも注目を浴びた理由は,ハテナが上記の光合成生物化の過程の内,段階1と段階2の間に位置する生き物と考えられるからです。ハテナについては既に多くの総説があるのでここではこれ以上は解説しません。 | ||||
| Endosymbiotic gene transfer (EGT) | ||||
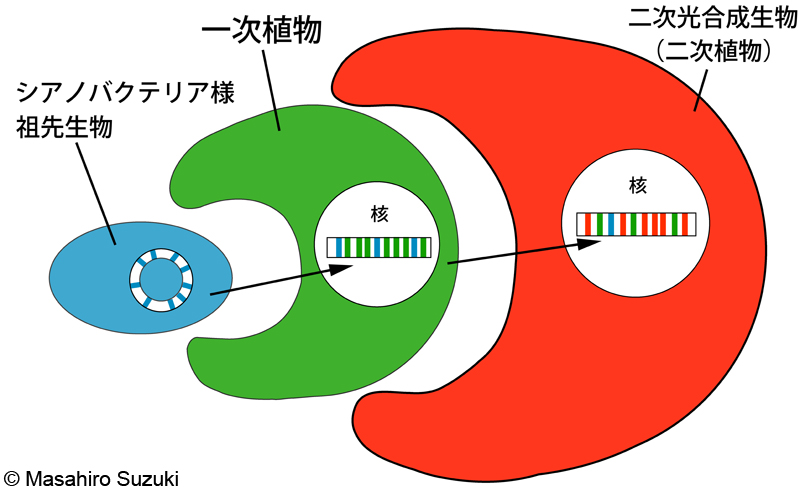
| 細胞内共生によって,共生体の遺伝子が宿主の核に移動する現象です。一次植物と二次光合成生物を用いた分子系統解析やゲノムを比較する際に留意しなければならない現象で,解析に用いた遺伝子がEGTに由来しているのか,宿主が独自に獲得した遺伝子なのかを見極める必要があります。 | ||||
 |
||||
| EGTの模式図:シアノバクテリア様の祖先生物の持っていた遺伝子(青)が細胞内共生(一次細胞内共生)によって,一次植物の核に移動します。一次植物の遺伝子(緑)と一次植物の核にコードされていたシアノバクテリア様祖先生物の遺伝子(青)が細胞内共生(二次細胞内共生)によって二次光合成生物(二次植物)の核に移動します。赤は二次光合成生物の核にコードされた遺伝子。EGTによって一次植物と二次光合成生物の核をコードする遺伝子はモザイク状になっている点がポイントです。*共生体の遺伝子が全て宿主の核に保存されるわけではありません。 | ||||
| 色素体タンパク質の輸送経路 | ||||
| 色素体のタンパク質は色素体自身が持つ遺伝子にもコードされていますが,その多くは核にコードされています(核コード色素体タンパク質遺伝子)。核コード色素体タンパク質遺伝子は翻訳された後,そのタンパク質を色素体まで輸送しなければなりません。核コード色素体タンパク質がどのような細胞内の経路をたどって,色素体まで届けられるのかは植物(藻類)グループによって異なります。また輸送の仕組み(例えると,荷物を送る際の荷札に相当する部分)もグループによって異なります。現在,いろいろな藻類グループを用いて色素体タンパク質輸送経路およびその機構の解明の研究が行われています。また,色素体タンパク質輸送経路を様々なクループで比較することで,色素体の成立機構やその進化を解明することにつながると期待されています。例えば,3枚の葉緑体包膜に包まれた色素体,4枚の膜に包まれている色素体,最外膜にリボソームが付着している色素緑体とではそれぞれのタンパク質輸送経路が異なります。近年,特に注目を集めているのは,祖先は同じ紅藻植物を共生体として持つ,アピコンプレクサと渦鞭毛藻類の色素体タンパク質輸送経路は4枚の膜に包まれるアピコンプレクサと3枚の膜に包まれる渦鞭毛藻類とで異なると考えられています。渦鞭毛類とクロメラの色素体タンパク質輸送経路の詳細が明らかになれば,色素体の成立機構やその進化の解明に大きな進展があると期待されます。 | ||||
| 「植物」という言葉 | ||||
| ここで述べたように,真核光合成生物には,我々が普段目にする緑色植物以外にも様々な生物がいます。Adl et al. (2005, 2012) などの現在の分類体系では,従来使われてきた「植物界(プランテ)」を用いず,一次植物である緑色植物,紅藻植物,灰色植物を「スーパーグループ アーケプラスチダ」に分類しています。アーケプラスチダは「最初の色素体」という意味で、一次植物は、最初に色素体を獲得したグループと考えられることから「最初の植物」,「真の植物」という意味を込めて名付けられたものです。一次植物に含まれる生物の多くは,「植物界(プランテ)」のメンバーだったことから,「スーパーグループ プランテ」としても良かったと思いますが,あえて「プランテ」という言葉を用いなかったのは,一般的に浸透している「光合成生物=植物」という概念を嫌ったものと考えられます。
日本では,光合成を行う生物を全て「植物」と呼び,その内,水中に生息し,維管束を持たないものを「藻類」と呼んでいます。従って,緑色植物とは系統的に異なるワカメ・コンブなどの褐藻類や渦鞭毛藻類,ミドリムシなどに対しても黄色植物,渦鞭毛植物,ミドリムシ植物として「植物」を用いています。しかし,系統的に異なる生物を全て「植物」と呼んで良いのか、光合成生物に対する一般的な理解を深めるためには、「アーケプラスチダ」のように「植物」という呼び方を止めてはどうか、あるいは「黄色生物」,「渦鞭毛生物」のように一次植物以外の光合成生物に対して「植物」という呼び方を止めてはどうかなど,様々な葛藤,意見があります。光合成生物,「植物」の正しい理解のためにはどのような用語を用いるべきか,今後の大きな課題です。 |
||||
| おわりに | ||||
| 色素体/葉緑体の成立と多様性について紹介してきました。ここで紹介しているのはあくまでも概要です。新しく発表された知見や紹介しきれていない事柄について,これからも更新していきます。 | ||||
| 参考文献 | ||||
| Adl, S.M., Bass, D., Lane, C.E., Lukeš, J., Schoch, C.L., Smirnov, A., Agatha, S., Berney, C., Brown, M.W., Burki, F., Cárdenas, P., Cepicka, I., Chistyakova, L., Campo, J. del, Dunthorn, M., Edvardsen, B., Eglit, Y., Guillou, L., Hampl, V., Heiss, A., Hoppenrath, M., James, T.Y., Karnkowska, A., Karpov, S., Kim, E., Kolisko, M., Kudryavtsev, A., Lahr, D.J.G., Lara, E., Le Gall, L., Lynn, D.H., Mann, D.G., Massana, R., Mitchell, E.A.D., Morrow, C., Park, J.S., Pawlowski, J.W., Powell, M.J., Richter, D.J., Rueckert, S., Shadwick, L., Shimano, S., Spiegel, F.W., Torruella, G., Youssef, N., Zlatogursky, V. and Zhang, Q. 2019. Revisions to the classification, nomenclature, and diversity of eukaryotes. Journal of Eukaryotic Microbiology 66:4-119. | ||||
| Adl, S.M., Simpson, A.G., Lane, C.E., Lukeš, J., Bass, D., Bowser, S.S., Brown, M.W., Burki, F., Dunthorn, M., Hampl, V., Heiss, A., Hoppenrath, M., Lara, E., Le Gall, L., Lynn, D.H., McManus, H., Mitchell, E.A., Mozley-Stanridge, S.E., Parfrey, L.W., Pawlowski, J., Rueckert, S., Shadwick, L., Schoch, C.L., Smirnov, A., Spiegel, F.W. 2012. The revised classification of eukaryotes. Journal of Eukaryotic Microbiology 59:429-493. | ||||
| Baldauf, S.L. 2003. The Deep Roots of Eukaryotes. Science 300: 1703-1706. | ||||
| Bodył, A., Stiller, J.W., and Mackiewicz, P. 2009. Chromalveolate plastids: direct descent or multiple endosymbioses?. Trends in Ecology and Evolution 24: 119-121. | ||||
| 井上 勲 2007. 藻類30億年の自然史 藻類から見る生物進化・地球・環境 第2版. 東海大学出版会,神奈川(秦野),643 pp. | ||||
| Maruyama, S., Suzaki, T., Weber, A.P.M., Archibald, J.M. and Nozaki, H. 2011. Eukaryote-to-eukaryote gene transfer gives rise to genome mosaicism in euglenids. BMC Evolutionary Biology 11: 105. | ||||
| Moore, R. B. et al. 2008. A photosynthetic alveolate closely related to apicomplexan parasites. Nature 451: 959-963. | ||||
| 高野義人 2010. アルベオラータに見られる多様な生き方~特に渦鞭毛藻類について~. 植物科学の最前線 第1巻 pp. 11-15. | ||||
| 宮城島進也 2014. 葉緑体の分裂制御機構とその進化. 植物科学の最前線 第5巻 pp. 21-36. | ||||
| Strassert, J.F.H., Irisarri, I., Williams, T.A. and Burki, F. 2021. A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids. Nature Communications 12: 1879. | ||||
| 山口晴代 2008. クレプトクロロプラストを持つ原生生物、特に渦鞭毛藻について. 原生動物学雑誌 第41 巻 第1 号 pp. 9-13. | ||||
| Yang, Y., Maruyama, S., Sekimoto, H., Sakayama, H. and Nozaki, H. 2011. An extended phylogenetic analysis reveals ancient origin of "non-green" phosphoribulokinase genes from two lineages of "green" secondary photosynthetic eukaryotes: Euglenophyta and Chlorarachniophyta. BMC Research Notes 4: 330. | ||||
| >藻類・プロチストの分類と解説 | ||||
| 「生きもの好きの語る自然誌」のトップに戻る | ||||
| © 2010 Masahiro Suzuki & Shuhei Ota | ||||