| ワカメ Undaria pinnatifida | ||||
| 作成者:鈴木雅大 作成日:2011年1月27日(2023年10月15日更新) | ||||
| ワカメ(若布,古語) | ||||
| Undaria pinnatifida (Harvey) Suringar 1873: 77. | ||||
| 黄藻植物(オクロ植物)門(Phylum Ochrophyta),クリシスタ(Chrysista),褐藻綱(Class Phaeophyceae),ヒバマタ亜綱(Subclass Fucophycidae),コンブ目(Order Laminariales),チガイソ科(Family Alariaceae),ワカメ属(Genus Undaria) | ||||
| *1. 吉田(1998)「新日本海藻誌」における分類体系:緑藻綱(Class Chlorophyceae),褐藻綱(Class Phaeophyceae),コンブ目(Order Laminariales),チガイソ科(Family Alariaceae),ワカメ属(Genus Undaria) | ||||
| *2. 吉田ら(2015)「日本産海藻目録(2015年改訂版)」における分類体系:褐藻綱(Class Phaeophyceae),コンブ目(Order Laminariales),チガイソ科(Family Alariaceae),ワカメ属(Genus Undaria) | ||||
| 掲載情報 | ||||
| 岡村 1902: 128; 1915: 273. Pl. 11, Figs 1, 2; 1926: 118. Pl. 231. Pl. 235, Figs 1-10; 1936: 281. Fig. 157; 遠藤 1911: 377-402. Figs 120-125; 川嶋 1989: 178. Pl. 57. Pl. 58, Figs 50, 51; 舘脇 1993: 136, 137. Fig. 68; 吉田 1998: 334, 335; Casas 2020: 83-86. Figs 1-9. | ||||
| Basionym | ||||
| Alaria pinnatifida Harvey 1860: 329; Suringar 1870: 24. Pl. 10. | ||||
| Homotypic synonym | ||||
| Ulopteryx pinnatifida (Harvey) Kjellman in Kjellman & Petersen 1885: 275; 岡村 1890: 45-48. Pl. 2. | ||||
| Heterotypic synonyms | ||||
| Alaria amplexicaulis G. Martens 1868: 114. | ||||
| Undaria pinnatifida var. vulgaris Suringar 1873: 77. Pl. 26 (V). | ||||
| Undaria pinnatifida [var. vulgaris] f. subflabellata Suringar 1873: Pl. 27 (VI). nom. nud. [裸名] | ||||
| Undaria pinnatifida [var. vulgaris] f. elliptica Suringar 1873: Pl. 28 (VII). nom. nud. [裸名] | ||||
| ナルトワカメ Undaria pinnatifida var. elongata Suringar 1873: 77. Pl. 29 (VIII). | ||||
| アオワカメ Laminaria peterseniana Kjellman in Kjellman & Petersen 1885: 267. Pl. 10, Figs 2, 3; 遠藤 1911: 370. Fig. 116. | ||||
| ≡ Undariopsis peterseniana (Kjellman) Miyabe & Okamura in 岡村 1902: 128. | ||||
| ≡ Undaria peterseniana (Kjellman) Okamura 1915: 275. Pl. 11, Figs 4-6; 1926: 129. Pl. 233; 1936: 283. Fig. 158; 川嶋 1989: 188. Pls 60-62, Fig. 53; 吉田 1998: 333-334; 宮田 2023: 487. | ||||
| ≡ Undariella peterseniana (Kjellman) Y.P. Lee 1998: 421. Figs 1-3. nom. illeg. [非合法名] | ||||
| ナンブワカメ Undaria pinnatifida var. distans Miyabe & Okamura in 岡村 1902: 128. | ||||
| ≡ ナンブワカメ Undaria distans (Miyabe & Okamura) Miyabe & Okamura in 宮部 1902: 57. Pl. 26. | ||||
| ≡ ナンブワカメ Undaria pinnatifida f. distans (Miyabe & Okamura) Yendo 1911: 708. Pl. 55, Figs 32-35. | ||||
| ヒロメ Hirome undarioides Yendo 1903: 99. Pl. 2, Figs 1, 2. Pl. 3, Figs 4-9; 1911: 375. Fig. 119; Yoshida 1991: 2. Pl. 9a. | ||||
| ≡ Undaria undarioides (Yendo) Okamura 1915: 274; 1926: 120. Pl. 232; 1936: 282; 川嶋 1989: 183. Pl. 59, Fig. 52; 吉田 1998: 335. Pl. 2-26, Fig. D. Pl. 2-27, Fig. D. | ||||
| ナルトワカメ Undaria pinnatifida f. narutensis Yendo 1911: 708. | ||||
| Undaria crenata Y.P. Lee & J.T. Yoon 1998: 438. Figs 7-10. | ||||
| Type locality: 静岡県 下田市 | ||||
| Holotype specimen: TCD (Trinity College, Dublin, Ireland) | ||||
分類に関するメモ:ワカメ(Undaria pinnatifida)には,種以下の分類群としてナンブワカメ(var. distans),var. elongata,var. vulgaris,f. elliptica,f. subflabellata,ナルトワカメ(f. narutensis)が記載されていますが,養殖ワカメのように密植状態では,ほとんどのワカメがナンブワカメ状になること(吉﨑 1999),Uwai et al. (2006) においても各ハプロタイプとこれらの種内分類群とが一致しないことから,本サイトでは種以下の分類群を採用しませんでした。Lee & Yoon (1998) は,Undaria crenata を記載しました。日本沿岸でもU. crenataに相当するものが採集されることがありますが,この種はワカメ,ヒロメ(U. undarioides),アオワカメ(U. peterseniana)のいずれかの雑種であるとの指摘があります。 Uwai et al. (2023) は,アオワカメ(Undaria peterseniana)とヒロメ(U. undarioides)をワカメ(U. pinnatifida)の異名同種(シノニム)としました。 |
||||
 |
||||
 |
||||
| 撮影地:兵庫県 淡路市 絵島(淡路島);撮影日:2015年4月22日;撮影者:鈴木雅大 | ||||
 |
||||
| 撮影地:兵庫県 豊岡市 竹野町 竹野 大浦湾;撮影日:2019年5月8日;撮影者:鈴木雅大 | ||||
 |
||||
 |
||||
| 撮影地:兵庫県 豊岡市 竹野町 竹野 大浦湾;撮影日:2015年5月8日;撮影者:鈴木雅大 | ||||
 |
||||
| 撮影地:兵庫県 淡路市 大磯(淡路島);撮影日:2017年3月29日;撮影者:鈴木雅大 | ||||
 |
||||
 |
||||
| 撮影地:青森県 下北郡 東通村 白糠;撮影日:2005年8月5日;撮影者:鈴木雅大 | ||||
| 海藻学者の憂鬱(?) | ||||
| ワカメの色をご存じでしょうか?本サイトで紹介している通り,ワカメは褐藻類の一種で,生きているときの色は黄色もしくは茶色(褐色)です。しかし,世間一般のイメージ及び各地で見られるワカメのイラストはどれも緑色です。味噌汁に入れるワカメや酢の物に入れるワカメはいずれも湯通ししたものなので,「ワカメは緑色」と思われるのも仕方ないかもしれません。著者も含め,海藻学者は各地で「ワカメは茶色(褐色)」と言い続けていますが,黄色や茶色(褐色)で描かれたワカメのイラストにはついぞお目にかかったことがありません。己の力不足を痛感します。いつか,「ワカメは褐色」が一般的になる日まで,海藻学者達の戦い(努力)は続きます。
・・・メディアで褐色のワカメを目にすることはないと思っていましたが,2020年10月に始まったあるアニメの第3話で,登場人物の頭に付いていたワカメ?は何と褐色でした。ワカメの形にはツッコミどころがあるものの,実写版で本物のワカメを使うならともかく,アニメで褐色のワカメを観る時が来るとは思ってもいませんでした。制作会社(ブリッジ)及び作画のこだわりに恐れ入りました。 さらに後日(2021年),子供がNintendo Switchの「あつ森」でワカメを採集して見せてくれたのですが,そのワカメは緑色ではなく,自然の色に近い黄褐色でした。当然緑色のワカメが出てくるものと思っていたため,驚きのあまり絶句しました。生きているワカメは緑色ではないということが,一般的に普及してきたのかもしれません。 |
||||
 |
||||
| 押し葉標本(採集地:静岡県 下田市 大浦海岸;採集日:2005年5月10日;採集者:鈴木雅大) | ||||
| ワカメのタイプ産地である下田で採集したワカメです。ワカメのような大型の海藻,特に「めかぶ」のように分厚く粘液に富んだ部分は,乾燥するのが難しく,ホルマリン固定していないものは乾燥中にカビが生えることもあります。この標本は,熱湯にくぐらせた後,「めかぶ」の裏半分をハサミで切り取ってから,押したものです。 | ||||
 |
||||
| 押し葉標本(採集地:岩手県 下閉伊郡 山田町 田の浜荒神;採集日:2004年7月4日;採集者:鈴木雅大) | ||||
| 三陸のワカメはとても大きくなります。3 mを超えるものが磯に普通に生えています。この標本は,150 cm位のワカメを台紙の表と裏を使って押し葉としたものです。茎が細長く典型的なナンブワカメ型です。ワカメの茎の長さは生育状態によって変わることから,分類学的にはナンブワカメ(Undaria pinnatifida var. distans)とナルトワカメ(U. pinnatifida f. narutensis)の区別はしません。ただし,これらの変種・品種は,歯応えや味に違いがあるため,流通の上では区別されます。また,産地によってブランド化しているところもあります。論文等で学名を用いる時には注意が必要ですが,普段の生活において,大きな問題はないと考えています。 | ||||
 |
||||
| 押し葉標本(採集地:新潟県 佐渡郡 真野町 滝脇(現 佐渡市 滝脇);採集日:2003年2月28日;採集者:鈴木雅大) | ||||
 |
 |
|||
| 幼体の押し葉標本(採集地:北海道 函館市 住吉;採集日:2012年4月7日;採集者:鈴木雅大) | ||||
| 紅藻ダルス(Devaleraea inkyuleei)を採集していたところ,紛れ込んできたものです。うっかり間違えそうになりましたが,かすかに見える中肋と羽状に生じた裂片からワカメの幼体だと気が付きました。 | ||||
 |
 |
|||
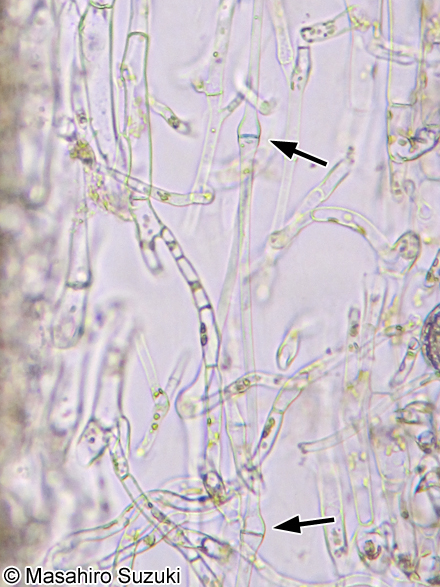
| 葉状部の縦断面 ラッパ状細胞(矢頭)が見られます。維管束植物の師管に良く似ています。 | ||||
| 胞子葉(めかぶ)の遊走子嚢から泳ぎ出した遊走子です。 | ||||
 |
||||
 |
||||
| 沖出し養殖の様子(撮影地:三重県 鳥羽市;撮影日:2014年1月21日;撮影者:鈴木雅大) | ||||
 |
||||
 |
||||
| 撮影地:新潟県 両津市 見立(現 佐渡市 見立);撮影日:2003年3月6日;撮影者:鈴木雅大 | ||||
| 養殖ワカメを採集(収穫)し,戻って来たばかりの船です。密植状態で養殖されたワカメは,どれも茎が長く,ナンブワカメ型になります。 | ||||
| ワカメ,ヒロメ,アオワカメは同じ種類? | ||||
| Uwai et al. (2006) は,ミトコンドリアにコードされるcox3遺伝子を用いた遺伝子解析の結果,ワカメ(Undaria pinnatidida),ヒロメ(U. undarioides),アオワカメ(U. peterseniana)は同種であると述べています。また,Saito (1972) は,交雑実験により3種はどの組み合わせでも雑種第一代(F)を作ること,ワカメとヒロメでは雑種第二代(F)まで作ることを報じています。自然界でもワカメとヒロメあるいはアオワカメとの雑種と考えられる個体がしばしば採集されており,Lee & Yoon (1998)が記載したUndaria crenataは,ワカメ,ヒロメ,アオワカメのいずれかの雑種であると考えられています(注)。Uwai et al. (2023) は,アオワカメ(Undaria peterseniana)とヒロメ(U. undarioides)をワカメ(U. pinnatifida)の異名同種(シノニム)としました。
注.Undaria crenataとワカメとの関係についてはっきりと論じた文献はありませんが,Casas (2020) は,吉田忠生博士の私信(T.Yoshida (in litt.))として,U. crenataがワカメ類の雑種の可能性があると述べています。 この状況は,マコンブ(Saccharina japonica var. japonica)とオニコンブ(S. japonica var. diabolica),リシリコンブ(S. japonica var. longipedalis),ホソメコンブ(S. japonica var. religiosa)の関係に良く似ていると思います。マコンブの場合,遺伝子解析の結果を基に,それまで別種と考えられていたオニコンブ,リシリコンブ,ホソメコンブをマコンブと同種とし,それぞれを変種(variety)の階級で区別しました(Yotsukura et al. 2008)。当時は,コンブの仲間の多くがLaminaria属からSaccharina属に移され(Lane et al. 2006),リシリコンブやホソメコンブがマコンブの変種となるという日本産コンブ類の分類が激変した時期で,海藻の中でも水産業や私達の生活との関わりが強いコンブ類の分類が変わるということで,混乱や反発が大いに危惧されました。また,マコンブ1種に統一せず,各種を変種の階級として区別したのは,オニコンブ,リシリコンブ,ホソメコンブの和名を残すためのやむをえない処置であり,必ずしも系統関係や地理的な要因を反映したものではないとの批判も予想されました。それから数年以上が経過しましたが,著者の予想に反し,懸念された混乱や反発はほとんど無かったように思います。また,その後行われた遺伝子解析において,各変種は交雑があるものの,それぞれの変種の集団が遺伝的にある程度のまとまりを維持していることから(Yotsukura et al. 2016),変種としての扱いも分類学的に妥当なものであったようです。 マコンブの前例をふまえれば,ワカメ,ヒロメ,アオワカメの場合も,「種」はワカメ1種とし,ヒロメとアオワカメをワカメの種内分類群として区別するのが適当かもしれません。ワカメ,ヒロメ,アオワカメの場合,マコンブと違い,それぞれの集団が遺伝的にはっきりと区別されず,ワカメ型,ヒロメ型,アオワカメ型のサンプル(ハプロタイプ)が系統内で複雑に入り組んでいます(Uwai et al. 2006, 2023)。従って,仮に種内分類群を認めるのであれば,品種(forma)の階級で区別するのが適当ではないかと思います。 |
||||
| アオワカメ(Undaria peterseniana)に相当する個体 | ||||
 |
||||
 |
||||
 |
||||
 |
||||
| 撮影地:千葉県 いすみ市 大原漁港;撮影日:2019年8月2日;撮影者:鈴木雅大 | ||||
| 千葉県いすみ市大原沖の器械根と呼ばれる漁場で行われたイセエビ漁の刺し網に掛かってきたアオワカメ(Undaria peterseniana)です。アオワカメは,千葉県レッドデータブック植物・菌類編 2023年改訂版において,重要保護生物(B)に選定されましたが(宮田 2023),ワカメと同種となったことで,選定の見直しが必要となりました。次回のレッドリスト,レッドデータブックの改訂において,「アオワカメ」をどのように扱うのが適当か検討中です。 | ||||
 |
||||
 |
||||
| 押し葉標本(採集地:静岡県 下田市 白浜海岸;採集日:2005年5月9日;採集者:鈴木雅大) | ||||
| アオワカメ(Undaria peterseniana)に相当する押し葉標本です。アオワカメは,ワカメ(U. pinnatifida),ヒロメ(U. undarioides)よりも深いところ(水深10~20 m)に生育し,中肋を持ちません。このため,海中ではコンブのように海底を這うように生育しています。また,胞子葉(めかぶ)を作らず,遊走子は体の中央部に形成します。これらの特徴から,形態的にはワカメ,ヒロメと区別されるため,ワカメと同種とされた際は大変驚きました。 | ||||
| 参考文献 | ||||
| Casas, G.N. 2020. Taxonomy and morphologies of Undaria pinnatifida (Phaeophyceae, Laminariales) in Golfo Nuevo, Argentina. Cymbella 6: 82-87. | ||||
| Guiry, M.D. and Guiry, G.M. 2023. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. https://www.algaebase.org; searched on 15 October 2023. | ||||
| Harvey, W.H. 1860. Characters of new algae, chiefly from Japan and adjacent regions, collected by Charles Wright in the North Pacific Exploring Expedition under Captain James Rodgers. Proceedings of the American Academy of Arts and Sciences 4: 327-335. | ||||
| 川嶋昭二 1989. 日本産コンブ類図鑑.215 pp. 北日本海洋センター,札幌. | ||||
| Kjellman, F.R. and Petersen, J.V. 1885. Om Japans Laminariaceer. Vega-expeditionens vetenskapliga iakttagelser, Stokholm 4: 255-280. | ||||
| Lane, C.E., Mayes, C., Druehl, L.D. and Saunders, G.W. 2006. A multi-gene molecular investigation of the kelp (Laminariales, Phaeophyceae) supports substantial taxonomic re-organization. Journal of Phycology 42: 493-512. | ||||
| Lee, Y.-P. 1998. Undariella, a new genus of the Alariaceae (Laminariales, Phaeophyta). Algae 13: 419-426. | ||||
| Lee, Y.-P. and Yoon, J.T. 1998. Taxonomy and morphology of Undaria (Alariaceae, Phaeophyta) in Korea. Algae 13: 427-446. | ||||
| 宮部金吾 1902. 北海道水産調査報告 巻之三.昆布採取業.216 pp. 北海道廳殖民部水産課. | ||||
| 宮田昌彦 2023. アオワカメ.In: 千葉県希少生物及び外来生物に係るリスト作成検討会(編)千葉県の保護上重要な野生生物-千葉県レッドデータブック-植物・菌類編 2023年改訂版.p. 487. 千葉県環境生活部自然保護課,千葉. | ||||
| 岡村金太郎 1890. わかめの生殖法.植物学雑誌 4: 45-48. | ||||
| 岡村金太郎 1902. 日本藻類名彙.276 pp. 敬業社,東京. | ||||
| Okamura, K. 1915. Undaria and its species. Botanical Magazine, Tokyo 29: 266-278. | ||||
| 岡村金太郎 1926. 日本藻類圖譜 第5巻 第7集.東京.*自費出版 | ||||
| 岡村金太郎 1936. 日本海藻誌.964 pp. 内田老鶴圃,東京. | ||||
| Saito, Y. 1972. On the effects of environmental factors on morphological characteristics of Undaria pinnatifida and the breeding of hybrids in the same genus Undaria. In: Abbott, I.A. and Kurogi, M. eds. Contributions to the systematics of benthic marine algae of the North Pacific. pp. 117-132. Japanese Society of Phycology, Kobe. | ||||
| Suringar, W.F.R. 1870. Algae japonicae. 39 pp. Typis Heredum Loosjes, Harlemi. | ||||
| Suringar, W.F.R. 1873. Illustrationes des algues du Japon. Musée Botanique de Leide 1: 77-90. | ||||
| 舘脇正和 1993. Undaria pinnatifida (Harvey) Suringar(ワカメ).In: 堀 輝三(編) 藻類の生活史集成 第2巻 褐藻・紅藻類.pp. 136, 137. 内田老鶴圃,東京. | ||||
| Uwai, S., Arai, S., Morith, T. and Kawai, H. 2006. Genetic distinctiness and phylogenetic relationships among Undaria species (Lamariales, Phaeophyceae) based on mitochondrial cox3 gene sequences. Phycologial Research 55: 263-271. | ||||
| Uwai, S., Takagi, S., Sekiguchi, T., Emura, N., Morita, T., Kurashima, A. and Sato, Y. 2023. Inconsistency between morphological diversity and genetic structuring: proposal for one species of Undaria in Japan. Botanica Marina 66: 81-90. | ||||
| Yendo, K. 1903. Three new marine algae from Japan. Botanical Magazine, Tokyo 17: 99-104. | ||||
| 遠藤吉三郎 1911. 海産植物学.748 pp. 博文館,東京. | ||||
| Yendo, K. 1911. The development of Costaria, Undaria, and Laminaria. Annals of Botany 25: 691-715. | ||||
| Yoshida, T. 1991. Catalogue of the type specimens of algae preserved in the herbarium, Department of Botany, The University Museum, The University of Tokyo. The University Museum, The University of Tokyo Material Reports No. 24. 17 pp. 67 pls. | ||||
| 吉田忠生 1998. 新日本海藻誌.1222 pp. 内田老鶴圃,東京. | ||||
| 吉田忠生・鈴木雅大・吉永一男 2015. 日本産海藻目録(2015年改訂版).藻類 63: 129-189. | ||||
| 吉﨑 誠 1999. 日本一のワカメ展.10 pp. 鯨と海の科学館,岩手県山田町. | ||||
| 吉﨑 誠 2005. ナルトワカメの謎 ー形態と味の地理的変異ー.国立科学博物館ニュース 437: 10-11. | ||||
| Yotsukura, N., Kawashima, S., Kawai, T., Abe, T. and Dreuhl, L.D. 2008. A systematic re-examination of four Laminaria species: L. japonica, L. religiosa, L. ochotensis and L. diabolica. Journal of Japanese Botany 83: 165-176. | ||||
| Yotsukura, N., Maeda, T., Abe, T., Nakaoka, M. and Kawai, T. 2016. Genetic differences among varieties of Saccharina japonica in northern Japan as determined by AFLP and SSR analyses. Journal of Applied Phycology 28: 3043-3055. | ||||
| >写真で見る生物の系統と分類 >真核生物ドメイン >S A R >ストラメノパイル >黄藻植物(オクロ植物)門 >褐藻綱 >コンブ目 | ||||
| >日本産海藻リスト >黄藻植物門 >褐藻綱 >ヒバマタ亜綱 >コンブ目 >チガイソ科 >ワカメ属 >ワカメ | ||||
| 「生きもの好きの語る自然誌」のトップに戻る | ||||
| © 2011 Masahiro Suzuki | ||||