2024年4月20日 カラタネオガタマ(被子植物,モクレン科),ツルソバ(被子植物,イヌタデ連),ルリチュウレンジ(昆虫,ミフシハバチ科)を公開しました。
2024年4月2日 青木政臣氏からヘビ類,カエル類の写真 9種を提供して頂き,ツシマアカガエルとチョウセンヤマアカガエル(カエル類,アカガエル科)を公開しました。
2024年3月30日 ゲンゲ(レンゲソウ)(被子植物,マメ亜科)を公開しました。
2024年3月13日 Kurita & Toda (2024) に従い,「クロイワトカゲモドキ」をヤンバルトカゲモドキ(爬虫類,トカゲモドキ科)に変更し,オビトカゲモドキ(爬虫類,トカゲモドキ科)の学名を変更しました。
2023年11月23日 Guiry & Frödén (2023) に従い,マリモ(緑藻類,アオミソウ科)の学名を変更しました。
2023年10月26日 アオヤガラ(条鰭類,ヤガラ科),クロガヤ(刺胞動物,アグラオフェニア科),ウミシダの1種(棘皮動物,ウミシダ目)を公開しました。
2023年9月24日 クサカゲロウの仲間の幼虫(昆虫,クサカゲロウ科)を公開しました。
2023年9月18日 Achnanthes sp.(珪藻類,ツメケイソウ科),Nitzschia sp.(珪藻類,クサリケイソウ科),Auricula sp.(珪藻類,ミミタブケイソウ科),Triceratium sp.(珪藻類,ミツカドケイソウ科),ウロコツツガタケイソウ(珪藻類,ツツガタケイソウ科)を公開しました。
2023年8月31日 底生性繊毛虫(繊毛虫類),プロトゾエア幼生(甲殻類,クルマエビ亜目),ハナオイアツバ(昆虫,ヤガ科)を公開しました。
2023年8月18日 Prorocentrum sp.(底生)(渦鞭毛藻類,フタヒゲムシ科),Pleurosigma sp.(底生)(珪藻類,メガネケイソウ科),クロミャクチャタテ(昆虫,チャタテ科)を公開しました。
2023年8月7日 バテイラ(軟体動物,クボガイ科)を公開しました。
2023年7月29日 イボイワオウギガニ(甲殻類,イワオウギガニ科),ゴマフニナ(軟体動物,ゴマフニナ科),ホンナガウニ(棘皮動物,ナガウニ科),ハタンポの仲間(条鰭類,ハタンポ科)を公開しました。
2023年6月18日 Micrasterias hardyi(接合藻,チリモ科),アカヒトリ(昆虫,トモエガ科)を公開しました。
2023年6月10日 コブカニダマシ(甲殻類,カニダマシ科),オウギガニ科の1種(甲殻類,オウギガニ科),ゴマフビロードウミウシ(軟体動物,ツヅレウミウシ科)を公開しました。
2023年5月21日 チビホコリタケ(?)(担子菌類,ホコリタケ科),ニホンカワトンボ(昆虫,カワトンボ科),オオムラサキ(幼虫)(昆虫,タテハチョウ科),シロシタホタルガ(幼虫)(昆虫,マダラガ科),スカシカギバ(幼虫)(昆虫,カギバガ科),ギンラン(被子植物,ラン科,セッコク亜科),コウボウシバ(被子植物,カヤツリグサ科),セリバヒエンソウ(被子植物,キンポウゲ科)を公開しました。
2023年3月14日 サカタザメ(軟骨魚類,サカタザメ科),カゲロウの仲間の幼虫(昆虫,カゲロウ目),カワゲラの仲間の幼虫(昆虫,カワゲラ目)を公開しました。
2023年1月7日 ウスイロコノマチョウ(昆虫,タテハチョウ科),コンジンテナガエビ(甲殻類,テナガエビ科),ミナミイワガニ(甲殻類,イワガニ科),ヘリトリアオリガイ(軟体動物,マクガイ科),ツノテツレイシ(軟体動物,アッキガイ科),マダライモ(軟体動物,イモガイ科)を公開しました。
2022年12月3日 Pachymeniopsis gargiuloi(紅藻,ムカデノリ科)を公開しました。
2022年9月9日 Phacus gigas(ミドリムシ門,ウチワヒゲムシ科),Rhabdostyla sp.(繊毛虫門,エピスティリス科),ネコゼミジンコ属の仲間(節足動物門,ミジンコ科),オナガミジンコ属の1種(節足動物門,シダ科)を公開しました。
2022年9月4日 カリオヒラムシ(扁形動物門,カリオプラナ科),アマオブネ(軟体動物,アマオブネ科),チョウトンボ(昆虫,トンボ科),ブチヒゲカメムシ(昆虫,カメムシ科)を公開しました。
2022年8月7日 ウミホタル(甲殻類,ウミホタル科),ベッコウクモバチ(昆虫,クモバチ科),ヤカドツノガイ(軟体動物,ゾウゲツノガイ科),スダレガイ(軟体動物,マルスダレガイ科),ハスノハカシパン(棘皮動物,ヨウミャクカシパン科),オカメブンブク(棘皮動物,ヒラタブンブク科)を公開しました。
2022年7月29日 ニホンイノシシ(哺乳類,イノシシ科)を公開しました。
2022年6月12日 ジュズモ属の1種(緑藻,シオグサ科)を公開しました。
2022年6月5日 キクスズメ(軟体動物,スズメガイ科),シロスジカミキリ(昆虫,カミキリムシ科),ヤケヤスデ(ヤスデ綱,ヤケヤスデ科)を公開しました。
2022年5月13日 オオカワヂシャ(被子植物,オオバコ科)を公開しました。
2022年5月1日 ヤセウツボ(被子植物,ハマウツボ科)を公開しました。
2022年4月16日 アブラコウモリ(哺乳類,ヒナコウモリ科)を公開しました。
2022年4月3日 オキカザシグサ(紅藻,ランゲリア科),ミドリハコベ(被子植物,ナデシコ科)を公開しました。
2022年2月6日 コウゼンジカワモズク(紅藻,カワモズク科)を公開しました。
2021年12月2日 エビイロカメムシ(昆虫,カメムシ科),オオハナアブ(昆虫,ハナアブ科),ナシイラガ(昆虫,イラガ科),フサコケムシの1種(外肛動物),タンキリマメ(被子植物,マメ科,インゲン連)を公開しました。
2021年10月5日 イワアミジ(褐藻,アミジグサ科)を公開しました。
2021年9月28日 コミスジ(昆虫,タテハチョウ科),オキナワカギバ(昆虫,カギバガ科)を公開しました。
2021年9月5日 シンカイヒメダルス(紅藻,マサゴシバリ科),トゲスズオビムシ(渦鞭毛藻,クリプトペリディニウム科),Coolia sp.(渦鞭毛藻,ピロキスチス科),ヒョウタンケイソウ(珪藻,ヒョウタンケイソウ科),エボシガイ(甲殻類,エボシガイ科),アカスジシロコケガ(昆虫,トモエガ科)を公開しました。
2021年8月15日 イトキヌゲ(紅藻,ランゲリア科),オオクワガタ(昆虫,クワガタムシ科)を公開しました。
2021年7月21日 ヤツダカワモズク(紅藻,カワモズク科)を公開しました。
2021年6月26日 Dinophysis sp.(渦鞭毛藻,カンムリムシ科),ヒラタオビムシ(渦鞭毛藻,ピロキスチス科),ヒラトゲガニ(甲殻類,イボトゲガニ科),オウギガニ(甲殻類,オウギガニ科),スベスベオウギガニ(甲殻類,スベスベオウギガニ科),ホソバハラアカアオシャク(昆虫,シャクガ科),ゴモクムシ亜科の1種(昆虫,オサムシ科),ハマエンドウ(被子植物,マメ科)を公開しました。
2021年5月15日 オジロアシナガゾウムシ(昆虫,ゾウムシ科),クロコガネ(昆虫,コガネムシ科),ベニカミキリ(昆虫,カミキリムシ科),カイコ(昆虫,カイコガ科),キショウブ(被子植物,アヤメ科)を公開しました。
2021年4月18日 Colaconema codicola(紅藻,コラコネマ科),セトウチフジマツモ(紅藻,フジマツモ科)を公開しました。
2021年3月31日 ミゾオキツノリ(紅藻,オキツノリ科)を公開しました。
2021年3月21日 ハダカモウミウシ科の1種(軟体動物,嚢舌目)を公開しました。
2021年3月11日 マイマイカブリ(昆虫,オサムシ科)を公開しました。
2021年3月9日 マゲカバノリ(紅藻,オゴノリ科)を公開しました。
2021年3月6日 ヒロバトガリエダシャク(昆虫,シャクガ科)を公開しました。
2020年12月8日 ヨナグニソウ,Yonagunia palmata,Y. taiwani-borealis(紅藻,ムカデノリ科)を公開しました。
2020年11月22日 Chondracanthus saundersii(紅藻,スギノリ科),Schottera koreana(紅藻,オキツノリ科),タチシノブ(シダ植物,イノモトソウ科),ミツデウラボシ(シダ植物,ウラボシ科),ノガリヤス(被子植物,イチゴツナギ連),ヤブマオ(被子植物,イラクサ科),ハナカタバミ(被子植物,カタバミ科)を掲載しました。
2020年11月14日 タテジマフジツボ(カイアシ・フジツボ類,フジツボ科),ウラギンシジミ(昆虫,シジミチョウ科),ツマジロエダシャク(昆虫,シャクガ科),オオクロバエ(昆虫,クロバエ科),カニクサ(シダ植物,カニクサ科)を追加しました。
2020年10月4日 カワトンボ科の幼虫,ホソミオツネントンボ(昆虫,アオイトトンボ科),クビカクシナガクチキムシ(昆虫,クビナガムシ科),マドガガンボ(昆虫,ガガンボ科),ミヤマカラスアゲハ(昆虫,アゲハチョウ科),オオアヤシャク(昆虫,シャクガ科),スジモンヒトリ(昆虫,トモエガ科),ウスベリケンモン(昆虫,ヤガ科)を追加・修正しました。
2020年9月29日 トウワタ(被子植物,キョウチクトウ科),キョウチクトウアブラムシ(昆虫,アブラムシ科)を追加しました。
2020年9月13, 16日 ムシクイ属の1種の幼鳥(鳥類,ムシクイ科),エゾギクトリバ(昆虫,トリバガ科),ヒナタイノコヅチ(被子植物,ヒユ科),スペアミントの1種(被子植物,シソ科,イヌハッカ亜科)を追加しました。
2020年8月21, 25日 シオミズツボワムシ(輪形動物,ツボワムシ科),ヒサゴクサキリ(昆虫,キリギリス科)を追加しました。
2020年8月16日 青木政臣氏からヘビ類,カエル類の写真 13種!を提供して頂き,ホルストガエル,オットンガエル,アマミハナサキガエル,アマミイシカワガエル(カエル類,アカガエル科),ガラスヒバァ(ヘビ類,ナミヘビ科)を追加しました。
2020年8月3, 5日 ヒメエグリバの幼虫(昆虫,ヤガ科),クロオビシロフタオ(昆虫,ツバメガ科)を追加しました。
2020年8月1日 ゴカクスケオビムシ(?)(渦鞭毛藻,スケオビムシ科),ヒドロクラゲ類の稚クラゲ(刺胞動物,アントアテカータ目),オベリアクラゲの仲間(刺胞動物,レプトテカータ目),ゴカイ科の幼生(環形動物,ゴカイ科),スピオ科の幼生(環形動物,スピオ目),舌殻類の幼生(腕足動物)を追加しました。
2020年7月23, 29日 アキカラマツ(?)(被子植物,キンポウゲ科),ハマナタマメ(被子植物,マメ亜科),コアカソ,ムカゴイラクサ(被子植物,イラクサ科),スナビキソウ(被子植物,ムラサキ科),ミヤマコゴメグサ(被子植物,ハマウツボ科)を追加しました。
2020年7月18, 19日 ツメウミウシ(?)(軟体動物,オキウミウシ科),スズメガイダマシ(腕足動物,シャミセンガイ目),キイトトンボ(昆虫,イトトンボ科),クロアゲハ(昆虫,アゲハチョウ科),コテングタケモドキ,シロオニタケの仲間(担子菌類,テングタケ科),オニイグチ(?)(担子菌類,イグチ科),ケショウハツ(担子菌類,ベニタケ科),ヒトモトススキ(被子植物,カヤツリグサ科),マツバハルシャギク(被子植物,キク亜科)を追加しました。
2020年7月11日 ホシスミイボゴケ(?)(地衣類,ピンゴケ科),モエギトリハダゴケ(地衣類,トリハダゴケ科),コキイロウラベニタケ(担子菌類,イッポンシメジ科),タマキクラゲ(担子菌類,ヒメキクラゲ科)を追加しました。
2020年7月6日 モハネガヤ(刺胞動物,ハネガヤ科),エゾカサネカンザシ(環形動物,カンザシゴカイ科),クサイロアオガイ(軟体動物,コカモガイ科),クビレクロヅケ(軟体動物,ニシキウズ科),コシアキトンボ(昆虫,トンボ科),サトユミアシゴミムシダマシ(昆虫,ゴミムシダマシ科),キオビツチバチ(昆虫,ツチバチ科)を追加しました。
2020年6月27-29日 ツルヒゲゴカイ(?)(環形動物,ゴカイ科),ショウジョウトンボ(昆虫,トンボ科),モンクロベニコケガ(昆虫,トモエガ科),オオシロカネグモ(クモ類,アシナガグモ科),イヌクグ(被子植物,カヤツリグサ科),コクチナシ(被子植物,アカネ科)を追加しました。
2020年6月23, 25日 ウルメイワシ(条鰭類,ウルメイワシ科),ハマナデシコ(被子植物,ナデシコ科)を追加しました。
2020年6月13, 15, 18日 ソエエダナシオキツ(紅藻,オキツノリ科),シロシュモクザメ(軟骨魚類,シュモクザメ科),セスジナミシャク,ウンモンオオシロヒメシャク(昆虫,シャクガ科),ハイモンカマトリバ(昆虫,トリバガ科)を追加しました。
2020年6月6日 Ecklonia cava var. kuromeoides(褐藻,コンブ科),クリアナアキゾウムシ(昆虫,ゾウムシ科),ノグルミ(被子植物,クルミ科)を追加しました。
2020年5月24, 31日 キエダシャク,キイロトゲエダシャクの幼虫(昆虫,エダシャク亜科),アオフシラクモヨトウ(?)(昆虫,ヤガ科),ヒメヒオウギ(被子植物,アヤメ科),キツネノボタン(被子植物,キンポウゲ連),ハルザキヤマガラシ(被子植物,アブラナ科),ナガバタチツボスミレ(被子植物,スミレ科),オヤブジラミ(被子植物,セリ科),ノボロギク(被子植物,サワギク連),コバノガマズミ(被子植物,レンプクソウ科),コナスビ(被子植物,ヤブコウジ亜科),ヤエムグラ(被子植物,アカネ科),トウバナ(被子植物,イヌハッカ亜科),ヒメイワダレソウ,ヤナギハナガサ(被子植物,クマツヅラ科)を追加しました。
2020年5月12, 17日 ニシイワヒゲ(褐藻,カヤモノリ科),ホンドテン(哺乳類,イタチ科),シメ(鳥類,アトリ科),クロイロコウガイビル(?)(扁形動物,コウガイビル科),アサヒナカワトンボ(昆虫,カワトンボ科),アカサシガメ(昆虫,サシガメ科),シロヘリカメムシ(昆虫,カメムシ科),ヤコンオサムシ(?)(昆虫,オサムシ科),ヤマトシリアゲ(昆虫,シリアゲムシ科),ガザミグモ(クモ類,カニグモ科)を追加しました。
2020年5月2, 5日 ナミウスカヤモ(褐藻,カヤモノリ科),キジ(鳥類,キジ科),ベリルイソギンチャク(刺胞動物,ウメボシイソギンチャク科),ニワハンミョウ(昆虫,オサムシ科),フタテンオエダシャク(昆虫,エダシャク亜科),キバラモクメキリガの幼虫(昆虫,ヤガ科),ナツフジ(被子植物,マメ亜科),ウシオツメクサ(被子植物,ナデシコ科),ナガエツルノゲイトウ(被子植物,ヒユ科),ノゲシ(被子植物,タンポポ亜科),ゴマギ(被子植物,レンプクソウ科),カキドオシ(被子植物,イヌハッカ亜科),マルバアオダモ(被子植物,オリーブ連),ヒメキンギョソウ,フラサバソウ(被子植物,オオバコ科)を追加しました。
2020年4月18日 メスアカケバエ(昆虫,ケバエ科),イヌシデメフクレフシ,アカシデメムレマツカサフシ(フシダニ科)を追加しました。
2020年4月4, 5日 ソラマメヒゲナガアブラムシ(昆虫,アブラムシ科),オオシモフリスズメ(昆虫,スズメガ科),キクラゲ(担子菌類,キクラゲ科),コガネニカワタケ(担子菌類,シロキクラゲ科)を追加しました。
2020年3月28, 29日 シロハラ(鳥類,ヒタキ科),ヒゲジロハサミムシ(昆虫,ハサミムシ科),ヒメフンバエ(昆虫,フンバエ科),クヌギハケタマフシ(昆虫,タマバチ科),カシワマイマイの幼虫(昆虫,トモエガ科),チャミノガ(昆虫,ミノガ科),マエアカスカシノメイガ(昆虫,ツトガ科)、キゴケの仲間(地衣類,キゴケ科),ホコリタケの1種(担子菌類,ハラタケ科),ニクハリタケ属の1種(担子菌類,シワタケ科),アラゲカワラタケ(担子菌類,タマチョレイタケ科),ヤブソテツ(シダ植物,オシダ科),ミチタネツケバナ(被子植物,アブラナ科),ヒメスミレ(被子植物,スミレ科)を追加しました。
2020年2月9日 ヒドリガモ(鳥類,カモ科),ツグミ(鳥類,ヒタキ科)を追加しました。
2020年2月2日 ケゼニゴケ(蘚類,ケゼニゴケ科),ホシダ(シダ植物,ヒメシダ科),イノデ(シダ植物,オシダ科),タカサゴユリ(被子植物,ユリ科),ツタ(被子植物,ブドウ科)を追加しました。
2020年1月25, 26日 トビモンオオエダシャク(昆虫,エダシャク亜科),クロコブタケ(子嚢菌類,クロサイワイタケ科),ニセショウロ属の1種(担子菌類,ニセショウロ科),コスギゴケ(苔類,スギゴケ科),ウラジロ(シダ植物,ウラジロ科),ハマスゲ(被子植物,カヤツリグサ科),ミドリサンゴ(被子植物,トウダイグサ亜科),ヒメユズリハ(被子植物,ユズリハ科)を追加しました。
2020年1月12, 18日 ミユビシギ(?)(鳥類,シギ科),ルビーロウカイガラムシ(昆虫,カタカイガラムシ科),ミスジミバエ(昆虫,ミバエ科),すす病菌の1種(子嚢菌類),シュタケ(担子菌類,タマチョレイタケ科)を追加しました。
2019年12月7, 14日 ツブダイダイゴケ(?)(地衣類,ダイダイキノリ科),トラノオシダ(シダ植物,チャセンシダ科),イノモトソウ,ホウライシダ(シダ植物,イノモトソウ科)を追加しました。
2019年12月1, 3日 サンカクフジツボ(カイアシ・フジツボ類,フジツボ科),アツミダイダイゴケ(?)(地衣類,ダイダイキノリ科),オオフサモ(被子植物,アリノトウグサ科),ヒマワリヒヨドリ(被子植物,キク亜科),キバナキョウチクトウ(被子植物,キョウチクトウ科)を追加しました。
2019年11月12, 17日 イセリアカイガラムシ(昆虫,ワタフキカイガラムシ科),クロスズメバチ(昆虫,スズメバチ科),ヒメヤママユ(昆虫,ヤママユガ科),ホラシノブ(シダ植物,ホングウシダ科),ニセアカシア(被子植物,マメ亜科),コウヤボウキ(被子植物,キク科),ダキバアレチハナガサ(被子植物,クマツヅラ科),クコ(被子植物,ナス科)を追加しました。
2019年11月5, 7日 ヒカリゴケ(苔類,ヒカリゴケ科),ノアサガオ(被子植物,ヒルガオ科),ソラスズメダイ(条鰭類,スズメダイ科),ミヤマハナゴケ(地衣類,ハナゴケ科)を追加しました。
2020年10月19, 22日 ナガキントキ(紅藻,ムカデノリ科),メマツヨイグサ(被子植物,アカバナ科),ハキダメギク(被子植物,キク亜科),クマツヅラ(被子植物,クマツヅラ科)を追加しました。
2020年10月14日 Chondracanthus okamurae(紅藻,スギノリ科),ヒオドシグサ(紅藻,フジマツモ科),アワフキムシの仲間の泡(昆虫,アワフキムシ上科),ヨモギクキワタフシ(昆虫,タマバエ科)を追加しました。
2019年9月29日 掲載種リストを公開しました。
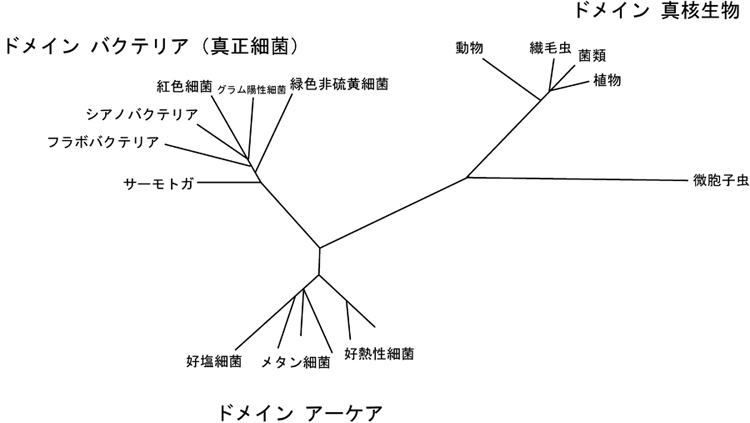
2019年6月1日 真核生物の分類・系統をAdl et al. (2019) に従って改訂しました。
2015年9月21日 赤尾正俊氏から昆虫類,キノコ類,カエル類,ヘビ類,軟体動物,陸上植物の写真 75種!を提供して頂きました。
2015年8月29日 湯浅智子氏から放散虫の写真 4種!を提供して頂きました。
2015年5月3日 Ruggiero et al. (2015) が発表した生物全体の分類体系を参考に,全体の分類体系を見直しました。
2013年12月25日 丸山健一郎氏からキノコ類の写真 53種!を提供して頂きました。
2013年5月24日 真核生物の分類・系統をAdl et al. (2012) に従って改訂しました。
2013年5月5日 青木政臣氏からヘビ類,トカゲ類,カメ類,カエル類の写真 34種!を提供して頂きました。全て野外で撮られた貴重な生態写真です。